Et av målene i BGE er å fylle i referansebiblioteket for europeiske arter. Det er et spesielt fokus på pollinatorer, ferskvannsarter, marine arter og arter viktige for miljøovervåkning, men materiale fra alle flercellede organismer er aktuelt. Det er utarbeidet en liste over dyr som mangler strekkoder i BOLD her. Velg gruppen du ønsker informasjon om og vent til tabellen lastes (det kan ta litt tid). Innsending av prøver og metadata er veldig likt det vi har gjort i NorBOL de siste årene og alle kostnader til sekvensering blir dekket av prosjektet. NTNU Vitenskapsmuseet og NHM i Oslo er partnere i prosjektet og kan hjelpe til med fasilitering. Ta gjerne kontakt med Torbjørn Ekrem om du ønsker mer informasjon om hva dette går ut på og hvordan du kan bidra.
Havforskningsinstituttet har en stor virksomhet rundt Afrika og opp til områdene rundt Sri Lanka og utenfor Myanmar (EAF-Nansen programmet). Her blir det blant annet samlet inn egg og larver som blir identifisering ved hjelp av DNA-strekkoding. Dette arbeidet har nettopp startet opp (våren 2019), men allerede nå er det identifisert over 150 arter og 2 arter som ikke tidligere er registrert i farvannet rundt Sri Lanka. Arbeidet utenfor Myanmar er bare i sin spede begynnelse med noen enkel egg prøver, men vil etter planen bli utvidet i 2020. Utfordringen med DNA-strekkodings arbeid i disse områdene er sannsynligvis en noe mangelfull database og dette er noe som vi må ha i tankene under arbeidet – skaffe oss prøver av voksne individ for verifisering og oppdatering av databasene. Kontaktpersoner: Padmini Dalpadado, Stamatina Isari.
Innsamling av plankton-nett. Foto Bjørn Krafft (c).
Havforskningsinstituttet har på oppdrag av Equinor Energy AS, Conoco Philips Scandinavia AS og Aker BP ASA analysert innsamlede egg og larver fra tre områder i Nordsjøen. Prosjektet ble iverksatt for om mulig å identifisere gyteområder for de økonomisk viktigste fiske artene i dette området med tanke på mulig seismiske aktivitet, men dataene som blir generert gir oss samtidig mye ny og oppdatert vitenskapelig kunnskap både om gyteområde og gytetidspunkt for ulike arter i Nordsjøen. Ved å sammenligne data fra tidligere survey i Nordsjøen og tilstøtende havområder kan dataene også brukes til å se på trender i vandringsmønster hos ulike arter, viktig kunnskap i forvaltningen av Nordsjøen. Til nå er det samlet inn over 2500 egg og larver som er identifisert til art v.h.a. DNA-strekkoding. Av de 141 fiskeartene som er kjent i Nordsjøen har vi til nå kunnet registrere 31 arter. Prosjektet er planlagt å avsluttes i august 2020. Kontaktperson: Bjørn Krafft
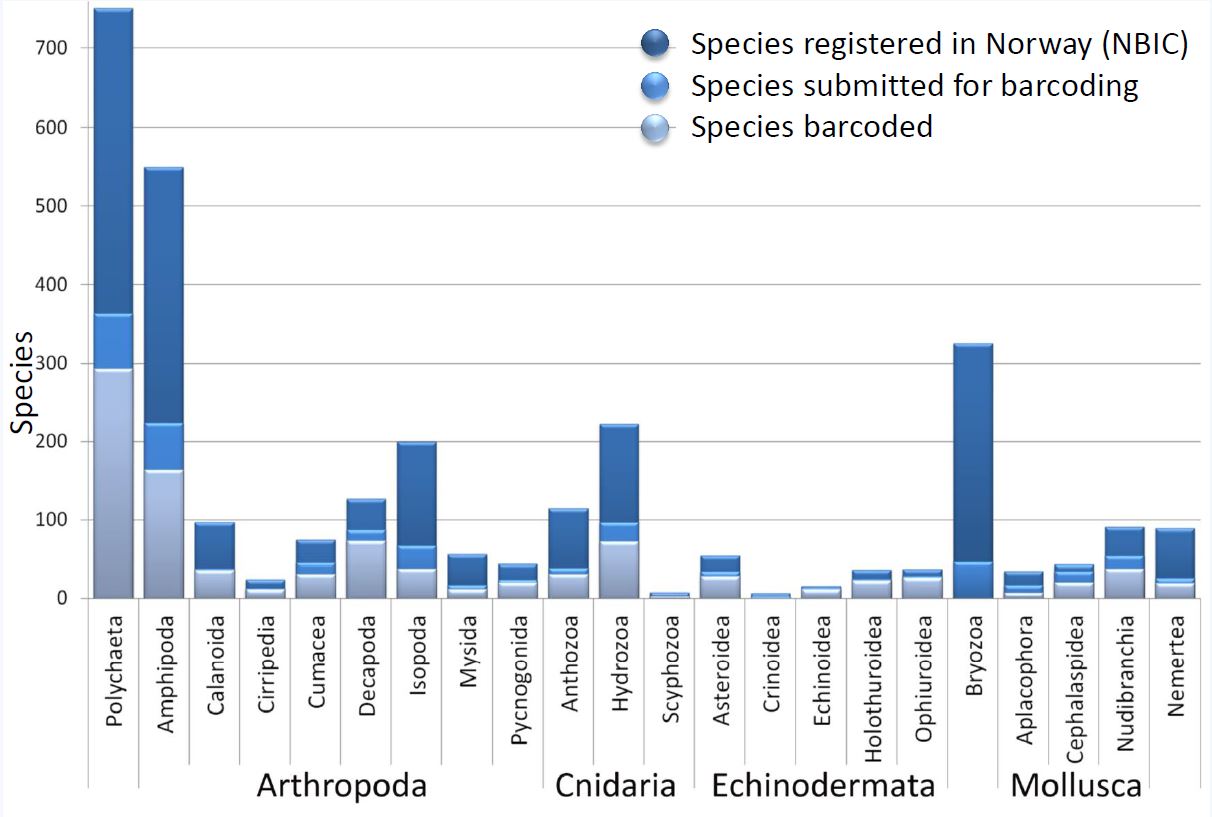
Disse resultatene, som ble presentert som poster 422 på den syvende IBOL-konferansen, gir et øyeblikksbilde på hvor langt vi har kommet i å bygge et kvalitetssikret strekkodebibliotek for marine virvelløse dyr (børstemark, pigghuder, krepsdyr, nesledyr, bløtdyr etc.) i fra Norge og tilstøtende områder (særlig Sverige, vi har samarbeidet mye med kollegaer i Gøteborg).
Før vi kunne sette i gang for alvor, måtte vi skaffe egnet materiale. Marint museumsmateriale er stort sett fiksert på formalin, som ødelegger DNA. Vi har derfor arrangert og deltatt på mange innsamlingstokt, og samarbeider også med store prosjekter som MAREANO og ulike miljøkartleggingsaktører for å skaffe prøver. Slik har vi de siste årene bygget opp store samlinger av spritfiksert materiale ved UM. Vi har også noen utfordringer med strekkodingsmetodikken; de såkalt «universelle» primerne som brukes i sekvenseringen har varierende suksessrate, og på enkelte dyregrupper vil det være nødvendig å inkludere andre markører (DNA-segmenter) enn standarden, som er cox1-segmentet.
Nå har vi – sammen med våre samarbeidspartnere – sendt inn over 8000 prøver til sekvensering. Av disse har vi fått sekvenser på 62 % av prøvene, og omkring 75 % av det vi kan kalle morfo-arter: dyr som vi enten kan identifisere til kjente arter basert på litteraturen, eller som vi midlertidig grupperer til «art A», «art B» etc. fordi vi ser at de skiller seg i fra de kjente artene.
Vi gjorde opp status for enkelte grupper i forhold til antallet kjente arter som finnes i Norge (basert på Artsdatabanken sine lister), og som du kan se er vi godt i gang på en del grupper – men det er fortsatt mye å ta tak i.
Det som ikke vises på denne figuren er de 2800 prøvene som vi ikke har fått strekkoder på med de gjeldene metodene, og de av prøvene som har fått strekkode, men som ikke har navn som korresponderer med Artsdatabanken sine lister over arter i Norge (dette kan skyldes f.eks. at arten ikke er registrert her, at den er ny for vitenskapen, eller at prøven vår ikke har blitt identifisert helt til art enda).
Her er to eksempler på observasjoner som bør følges opp med taksonomisk utredning:
Først ut er en stor sjøstjerne i fra slekten Diplopteraster. I fra Norge har tre individer av D. multipes blitt strekkodet. Når strekkodene for disse dyrene sammenlignes med andre i den store BOLD-basen så er de identiske med strekkoder som er produsert av andre marinbiologiske laboratorier, men som er identifisert med fem ulike artsnavn basert på morfologien.
Dette er verdt å se nærmere på – er de fem artene egentlig bare én vidt utbredt art, som vitenskapelige kulturer i ulike land har forstått som ulike arter, særegne for sin geografiske region? Eller er det den genetiske markøren vi har brukt som ikke klarer å skille mellom faktiske arter i denne gruppen? Slike spørsmål krever nærmere undersøkelser med taksonomiske studier, der det fysiske materialet som artsforståelsen er basert på kan studeres med kritisk blikk på den vitenskapelige litteraturen. I tillegg bør slike vurderinger av artsstatus helst ha data fra andre genetiske markører slik at både strekkoder og morfologiske karakterer kan vurderes i nytt lys. Noen ganger kan uoverensstemmende artsnavn for identiske strekkoder ganske enkelt skyldes menneskelig feil, – at den som navnsatte en strekkode tok feil i sin artsbestemmelse basert på morfologiske karakterer.
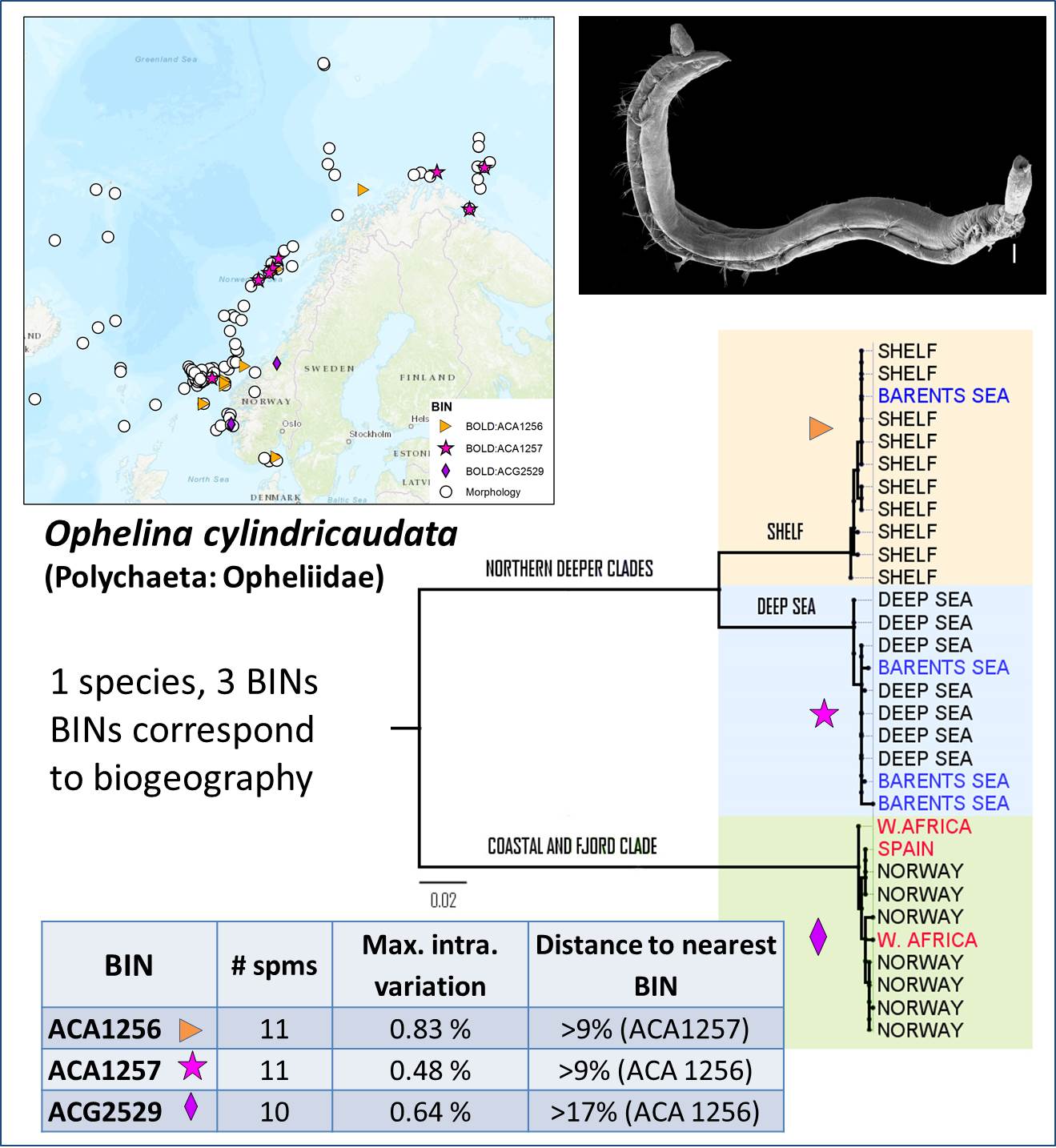
I det neste eksemplet har vi en antatt art med overraskende stor genetisk variasjon i det strekkodede materialet. Spørsmålet blir da om børstemarken Ophelina cylindricaudata i realiteten består av flere ulike arter. Det ser ut for at de genetiske gruppene som framkommer av strekkodedata kan kobles til ulike habitater som markene lever i.
Igjen viser det seg et behov for nærmere taksonomiske, økologiske og evolusjonsbiologiske studier. Etterhvert er det ikke lengre uvanlig at nærmere studier av såkalt kryptiske arter først «avsløres» med strekkoder og ved nærmere studier kan vise seg å også ha morfologiske kjennetegn som skiller dem fra sine nærmeste slektninger.
Tilsvarende problemer som disse to eksemplene finnes det mange av. De viser at det er langt igjen til en presis og korrekt forståelse av artsmangfoldet i marine miljøer. Men med strekkoding kan vi finne hvor problemene befinner seg og slik sett identifisere behovene for videre arbeid. Selvfølgelig må vi heller ikke glemme at kvalitetssikrede strekkoder også gir vidtrekkende muligheter for så vel biologer som andre profesjoner for å identifisere organismetyper med stor presisjon, enten de foreligger som levende, døde, eller som DNA-spor.
Katrine Kongshavn, Jon A. Kongsrud, Tom Alvestad, Endre Willassen, Universitetsmuseet i Bergen, avdeling for naturhistorie.
Disse resultatene ble presentert på 7th International Barcode of Life Conference, Sør-Afrika, 2017.
Ved å benytte NTNUs nettsider samtykker du i at vi kan etterlate informasjonskapsler i din nettleser.
By using NTNU's websites, you agree that we may leave cookies in your browser.
 English
English  Norwegian Bokmål
Norwegian Bokmål